The study of migration patterns in adult Pacific halibut spans the majority of the IPHC’s research history and has employed a variety of tagging techniques in an attempt to better understand numerous migratory processes that occur over multiple temporal and spatial scales. These range from long-distance migrations along the coastline that may cover thousands of kilometers and require years to complete; to offshore spawning migrations. The IPHC recently reviewed the last 100 years of migration research available for Pacific halibut, with a focus on tagging experiments and presented a conceptual model of seasonal migrations from larval to adult life history stages than spans the entire distribution range of the species in the North Pacific Ocean (Carpi et al. 2021). Specific details on the different types of adult migration of Pacific halibut can be found in the following sections.
Long-distance (ontogenic) adult migration
Over the course of the lifetime of Pacific halibut, average net migration among all individuals in the benthic phase (i.e., after settlement) is to the south and east. This must be true if the population is to maintain the same geographic distribution over time, because ocean currents transport the larvae to the north and west (for more on larvae, see: Larval distribution). If juveniles and adults did not migrate in the opposite direction, the entire population would eventually disappear from the eastern Pacific Ocean. This component of migration that occurs after settlement may be referred to as “contranatant”, meaning that it is in the opposite direction of (it counters) the drift that occurred as larvae. In general, long-distance movements that occur over an individual’s lifetime are known as “ontogenic” (or ontogenetic), meaning that they occur over the course of the development of the individual (i.e. during their ontogeny). For information regarding migration of juveniles, see: Juvenile migration. In this section can be found a brief survey of what we understand of adult migration; or more accurately, considering that maturity status (i.e. that which distinguishes a juvenile from an adult) is rarely known for any individual that is tagged and released, the migration of “large” Pacific halibut that are expected to be mature based on a broader understanding of maturity at size (for more on maturity, see: Reproductive assessment).
From Conventional Tagging
Over the course of the IPHC’s history, nearly 400,000 Pacific halibut of various sizes have been marked and released bearing various forms of conventional tags, generating more than 46,000 tag recoveries (Kaimmer 2000). Most of these fish were tagged in the Gulf of Alaska, including Canadian waters. Tagging of large Pacific halibut for the purposes of understanding their migration among grounds and regions was an early priority of the IPHC, comprising the Banks Tagging Program conducted from 1925-1940 (Thompson and Herrington 1930; Kaimmer 2000). These studies derived our fundamental understanding of the migration of Pacific halibut of exploitable sizes (Skud 1977): 1) the potential for individuals to move great distances, but for the majority of fish to be recaptured in the Regulatory Area in which they were tagged; 2) generally declining rates of annual migration with age and size; 3) a tendency for movement to be greater to the east and south than to the west and north, and; 4) very little movement of large individuals from the Gulf of Alaska into the Bering.
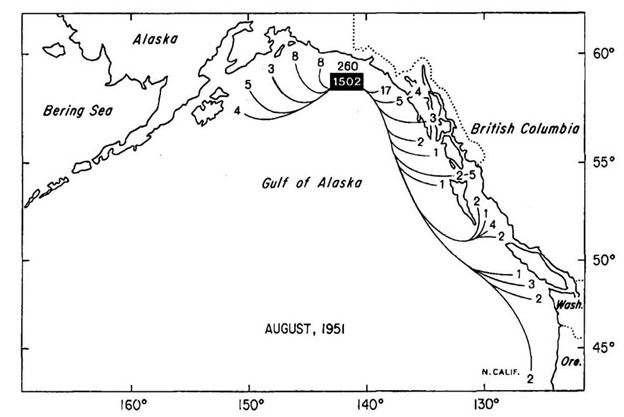
Figure 1. Distribution of recoveries from Pacific halibut tagged off Yakutat in August, 1951. Note the greater tendency for eastward than westward movement. From: Skud (1977), IPHC Sci. Rep. 63.
Modern tagging programs have sought to refine our understanding of adult movements and to quantify rates and identify specific behaviors, but this general model of dispersal remains robust.
From Passive Integrated Transponder (PIT) Tags
From 2003-2009 the IPHC conducted the largest single tag release and recovery experiment in its history to-date. Over a two-year period, approximately 67,000 Pacific halibut were tagged using Passive Integrated Transponder (PIT) tags: glass tags approximately the size of a grain of rice, that were implanted below the skin of the fish on their pre-opercular plate (i.e. on the side of the head). PIT tags contain a small coil, ferrite rod, and microchip, and when placed into a suitable electrical field are induced to emit an individually-coded signal. PIT tags were selected for this release because its primary intent was to analyze tag return rates in order to estimate regional exploitation rates within the longline fishery and to estimate natural mortality Tag data can only be reliably used for such analyses if the returns are not biased by factors such as regionally-variable reporting rates. Given that the PIT tags could not be seen by the fishers who recovered them, concerns of reporting bias were therefore eliminated. Instead of relying on fishers to report their recoveries, the IPHC established a tag-scanning program that was associated with its port sampling operations, and systematically monitored commercial landings at major ports in each IPHC Regulatory Area from 2003-2009.
Although the tagging protocol was “blind” to whether the tagged fish were adult or juvenile, by simply tagging individuals across all encountered sizes in proportion to their surveyed abundance, a large number of both demographics were presumably represented. Ultimately, robust estimation of exploitation and mortality rates proved impossible and displayed departures from assessment-based estimates in a regionally-trended manner likely due to regionally-variable violations of the underlying assumption that all of the tagged fish would mix quickly and thoroughly into the population. However, a wealth of information regarding migration was obtained (Webster et al. 2012). This included verification of the results of earlier conventional tagging studies (Dunlop et al. 1964, Bell 1967, Bell and Best 1968, Skud 1977) and analysis (Quinn et al. 1990) that – despite being prone to reporting issues – had indicated that contranatant migration in Pacific halibut continues into adulthood and at sizes that are recruited to the longline fishery. The new information also allowed for updated estimates of migration-by-size to be derived (Webster et al. 2012).
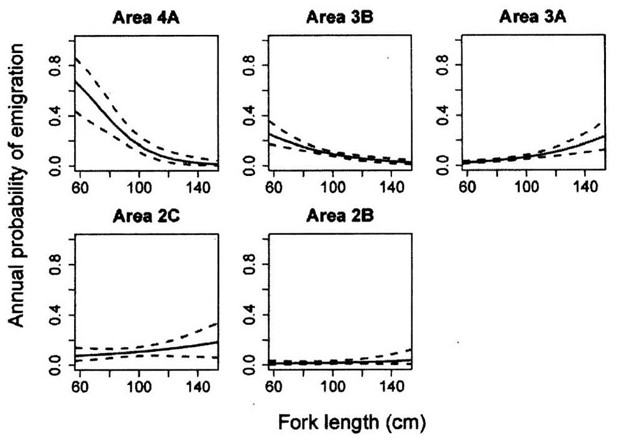
Figure 2. Estimated annual rates of emigration of Pacific halibut from IPHC Regulatory areas, according to fish length. Solid lines are fits from a tag-recovery model (for model details, see: Webster et al. (2012), Can. J. Fish. Aquat. Sci. 70: 642-653), with dashed lines representing 95% posterior credible intervals.
From pop-up archival transmitting (PAT) tags
The tagging programs described above have given us a solid foundation for understanding adult migration of Pacific halibut, but even the PIT tag results are constrained by biases that are inherent in all physical mark-recapture studies: one can only index the movement of fish away from or within areas where the tags were deployed; fish can only be observed to move to areas where there are recapture mechanisms (here: fisheries); and conclusions regarding the rate of exchange among areas may be biased by the relative probability of recapture for fish moving into each of the indexed areas. For example, the IPHC’s understanding of movement into and out of the eastern Bering Sea – especially to the north – is undoubtedly biased by the existence of a biologically-artificial boundary between US and Russian waters. The IPHC has rarely tagged fish in Russian waters; and the return of tags that are recaptured in Russia is quite unlikely. As such, we are necessarily prone to assume that the northeastern Bering Sea shelf edge is a net exporter of Pacific halibut (because rates of immigration from Russia are entirely unknown), and that fish that reside in this region do not disperse westward. Similarly, one of the results of the PIT tagging experiment was to provide strong evidence that the probability for tagged Pacific halibut to be recaptured is much smaller in the Aleutian Islands region than elsewhere. As such, even if the majority of fish tagged in the Aleutian Islands remain there, the tag data are prone to suggesting that most will migrate out of the region (because when they do, their chance of being recovered greatly increases); and rates of migration westward into the Aleutian Islands from other regions is prone to being substantially underestimated (because fish migrating in this direction are unlikely to be recaptured).
However, there is one type of tag that is not subject to these biases: Pop-up Archival Transmitting (PAT) tags. PAT tags are electronic tags that are attached to fish using a dart-and-tether mechanism; the dart is embedded into the fish, and the other end of the leader is attached to a release mechanism on the end of the tag. On a pre-programmed date, the mechanism releases the tag from the fish and the tag floats to the sea surface to emit radio signals that are received by the Advanced Research and Global Observation Satellite (ARGOS) system. The satellites use the tag’s signal to calculate a location in the same way that a vessel’s position is determined when an EPIRB (Emergency Position Indicating Radio Beacon) is activated. That position is then made available to the researcher. Because the entire ocean surface is monitored by these satellites, the fish’s position can be obtained without the need to recapture the fish. Also, if the tag is shed by the fish prior to the programmed date or is recaptured by an individual who chooses not to report the recovery, the tag will still broadcast upon surfacing and its position will still be reported to the researcher. The result is an ability to study dispersal without recapture or reporting biases. Still, PAT tags have their own set of limitations: they are large, so can’t be used on halibut smaller than ~50 cm in length; they cause drag, and so have the greater potential to affect fish behavior than do smaller tags, especially as fish size declines; they can be prone to shedding after long periods, and so deployments of greater than two years are not advisable; they are expensive, ranging from US$1500-$4000 depending upon configuration.
Given evidence for bias in observed PIT-tag recapture rates in the Bering Sea and along the Aleutian Islands, the IPHC conducted a study from 2008-2010 (Loher and Clark 2010) to more objectively study dispersal of exploitable stock in the region. The design focused on fish of all commercially-exploitable sizes (i.e. >82 cm fork length) – hence, presumably a mixture of late juvenile and adult females and mature males – distributing 107 PAT tags from just south of Unimak Pass to the continental shelf edge near the Russian border, and westward through the Aleutian Islands to Attu Island. The tags were programmed to report their positions 365 days after tagging. End positions were obtained for 93 of those fish (14 tags failed to report): positions for 66 of those were obtained the following late spring or summer, at or near one year after tagging; positions for 27 fish were obtained from fall of the tagging year to early spring of the following year. The primary observations were that: 1) dispersal occurred across the Russian border; 2) fish tagged west of Samalga Pass in the Aleutian Islands remained in the Aleutian Islands, and showed no evidence of crossing either Samalga or Amchitka Pass; 3) fish tagged in the Bering Sea showed no evidence of moving into the Gulf of Alaska; 4) fish that were tagged in IPHC Regulatory Area 4A south of Unimak Pass (i.e. in the far-western Gulf of Alaska) displayed high rates of eastward dispersal, into Areas 3 and 2. It must be cautioned that the small sample sizes likely prevented us from detecting migrations that occur at relatively small rates. However, it is notable that the general patterns are consistent with prior research, particularly the apparent discontinuity in mixing between the Bering Sea and Gulf of Alaska that was also apparent in the PIT tag data. The functional structure that was revealed in this experiment is consistent with the spatial structure that comprises the IPHC’s Areas as Fleets assessment model structure (Stewart and Martell 2015).
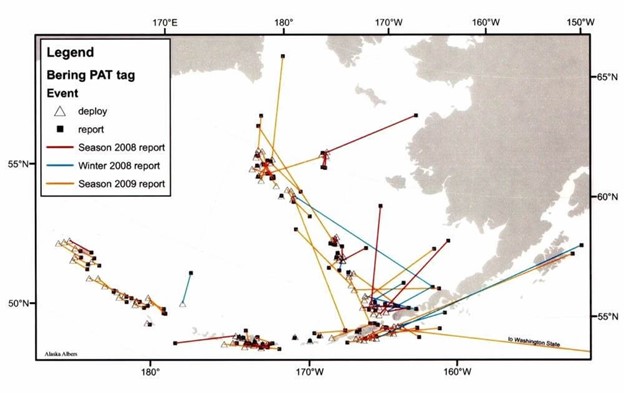
Figure 3. Deployment (open triangles) and reporting (solid squares) locations for PAT tags deployed on Pacific halibut ≥82 cm fork length during the summers of 2008 and 2009. Most notable are: dispersal northward across the USA-Russian border, and apparent discontinuities in (i.e. no evidence of) dispersal along an east-west axis across deep Aleutian passes or southward through Unimak Pass.
Seasonal migration
In addition to the long-term one-way migrations that Pacific halibut conduct as they age, adult Pacific halibut also undertake annual two-way migrations between their summer feeding grounds and winter spawning sites. Generally, Pacific halibut feed on the continental shelf and spawn in relatively deep waters (>200 m; Loher and Seitz 2008) along the continental slope, concentrated at submarine canyons from at least British Columbia to the southeast Bering Sea (St. Pierre 1984); and likely as far south as Washington State (Loher and Blood 2009), westward to Attu Island (Seitz et al. 2011), and northward to at least Middle Canyon (Seitz et al. 2011). These migrations may occur on two axes: roughly parallel to the coastline (i.e. alongshore); and onshore-offshore. The magnitude and timing of these migrations can have important implications for a management structure that employs a closed winter season to control effort and a summer survey to guide decisions regarding the spatial distribution of removals.
Alongshore migration, from conventional tagging
One of the IPHC’s first management actions was to impose upon the Pacific halibut longline fishery a winter closure that was intended as a form of effort limitation, wherein harvests were maintained within target levels by adjusting season length according to the fleet’s fishing power (Babcock et al. 1931). Although the Commission later concluded that the closed season did not result in stock conservation, the winter closure was maintained due to its “obvious beneficial economic effects” (Babcock et al. 1931) and it has persisted ever since. Seasonal management of the fishery has progressed through three eras, characterized by single or multiple summer fishing periods and different degrees of variability in season length. Single annual seasons were employed from 1925 to 1953, but decreased from nine months to one month as fleet power increased. Multiple fishing periods were invoked in 1954, after which the fleet continued to grow and the length of the season continued to contract, reaching its shortest length (36 hours) in the Bering Sea in 1991. The adoption of Individual Quota (IQ) management in 1991 in Canada and in 1995 in Alaska re-established a single extended summer fishery from Canada northward, with a roughly three-month winter closed period. In 1998, economic concerns prompted the fishing industry to formally request that the IPHC substantially extend the commercial season (Leaman and Clark 2000), and an interagency working group was convened to review issues associated with implementing that proposal (Gilroy and Sadorus 2000). It was concluded that extending commercial halibut harvests into the winter spawning season would present a number of challenges, one of which being the expectation for seasonal migration to cause substantial redistribution of exploitable stock among areas and generate uncertainty regarding the spatial effects of removals applied during winter (Leaman and Clark 2000).
In an attempt to estimate the magnitude of seasonal redistribution between Canadian waters (IPHC Regulatory Area 2B) and Alaskan waters to the north, an analysis was conducted using conventional mark-recapture tag data (Leaman et al. 2002). In the absence of a targeted fishery to provide sufficient winter recovery data, the analysis relied on tags that had been deployed between October and April (Skud 1977, St. Pierre 1984, Kaimmer 2000) and subsequently recovered in summer fisheries. Based on 2,620 tag returns, it was suggested that as much as 2/3 of spawning stock that is resident in Canadian waters during the summer should be expected to move to Alaskan waters during the winter and be vulnerable to out-of-area interception. However, it was cautioned that the results could be subject to spatial biases in tag recovery rate and that the estimates derived from the analyses should not be considered sufficiently precise to estimate the impacts of intercepting seasonally-migratory fish.
Alongshore migration, from PAT tags
In a desire to more accurately estimate seasonal movement of Pacific halibut spawning stock between Canadian and US waters, using fishery-independent location data obtained at the end of the winter closure period, a PAT-tagging experiment was conducted in 2008-2009 (Loher and Blood 2009). Seventy-eight fish ranging from 104-163 cm in fork length were tagged during the summer in the USA Pacific Northwest (Area 2A) and Canada (Area 2B). Based on the conservative assumption that all of the tagged fish were female and an understanding of regional maturity-at-size, it was estimated that roughly 80% of the tagged fish would be mature; a higher proportion if the tagged population included males. Tags were programmed to detach and report their locations on 1 February, 15 February, and 1 March, representing three potential season-opening dates that were incrementally earlier than the traditional date of 15 March. Fifty-seven tags generated position data on the specified dates. The results suggested that seasonal redistribution is likely to be considerably lower than estimated from conventional tag returns (Leaman et al. 2002): movement from Canadian to Alaskan waters was 7%; emigration from the USA Pacific Northwest to Canada was 30% and to Alaska 10%. However, the results could be subject to demographic bias because it was designed to investigate movement of female spawning biomass, and an unknown proportion of fish may have subjected themselves to undetected risk of interception by departing their tagging region but returning prior to their tag-reporting date.
A more-comprehensive suite of PAT-tagging experiments has been conducted coastwide that have been designed to yield a less-biased understanding of seasonal redistribution. In these studies, large Pacific halibut (possessing a high probability of being mature females) were tagged during the summer with tags programmed to detach and report in January, during the peak of the spawning season (Loher and Seitz 2008). In addition to the location data obtained on the pop-up date, the depth data recorded by the tag during the period at liberty and at-liberty position estimates derived from archived light data (see sections below) allow for each fish’s record to be evaluated for the possibility that it migrated offshore and returned to its tagging site prior to tag reporting. Experiments falling within this suite were initiated in 2002, with the most recent deployments occurring in 2017 and expected to produce results in 2018. The results of these studies through 2008 are summarized in Seitz et al. (2017) and are broadly consistent with the results of the annual dispersion experiment described above (Loher and Clark 2010): Pacific halibut that summer in the Aleutian Islands show a strong tendency to remain within their islands groups during the spawning season; dispersal along the eastern Bering Sea continental shelf edge appears common, while exchange of spawners between the Bering Sea and Gulf of Alaska is limited; and dispersal within the Gulf of Alaska and US Pacific Northwest is fairly extensive. Again, it is noteworthy that the spatial stock structure implied by these results is consistent with the structure of the IPHC’s Regulatory Areas as Fleets assessment model (Stewart and Martell 2015).
Onshore-offshore Migration and Spawning, from Electronic Archival Tags
Whereas conventional and PIT tags produce, at best, two data points per fish – fish location at tagging and recovery – electronic archival tags (including PAT tags and smaller fishery-recovery tags) can produce millions of data points per fish per year: tagging location, recovery location, temperatures experienced and depths visited every 30 seconds or less, and the potential for daily position estimates to be derived using light data (Seitz et al. 2006), geomagnetism (Kobayashi et al. 2008), and tidal-amplitude estimation (Hunter et al. 2004). Depth data obtained from electronic archival tags are ideal for identifying and characterizing the onshore-offshore component of seasonal migration, generally documented decades ago at the population level (St. Pierre 1984), and has comprised a component of most of the electronic archival tagging studies that the IPHC has conducted to-date. For Pacific halibut that undertake seasonal migrations, the annual depth profile is most-often characterized by a shallow summer phase followed by movement to deep water in the autumn, a secondary movement to an even deeper depth stratum in winter, and return to shallow water the following spring.
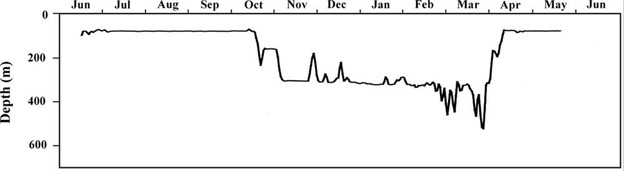
Figure 4. A typical seasonal depth profile for a likely-female Pacific halibut conducting an offshore spawning migration. Note the two depth strata that were visited during the winter. In this individual, a series of putative spawning rises was observed during late February and early March.
Additionally, examination of the detailed archival records has revealed a type of vertical migratory behavior that is conducted at much smaller scales: series of abrupt ascents, 100–200 m in magnitude, each of ~10 minutes in duration, conducted during mid-winter. In many cases, this behavior appears in the archival record during the fish’s movement to its deepest annual depths. This behavior is believed to represent active spawning in Pacific halibut (Seitz et al. 2005) because it closely parallels spawning behavior observed in other flatfish species (Moyer et al. 1985, Konstantinou and Shen 1995, Manabe and Shinomiya 2001, Carvahlo et al. 2003, Murphy et al. 2017), and is consistent with the fact that Pacific halibut are serial (batch) spawners (St. Pierre 1984). In other species that conduct these rises, the female joins its mate (or mates) on the seafloor before the rise begins, they ascend together to the apex of the rise where the eggs are released and fertilized, and the fish then descend to the bottom. This process of midwater egg fertilization is hypothesized to result in lower loss of eggs to predation than would occur near-bottom, as well as placing the eggs closer to the surface waters where the larvae will eventually drift and feed.
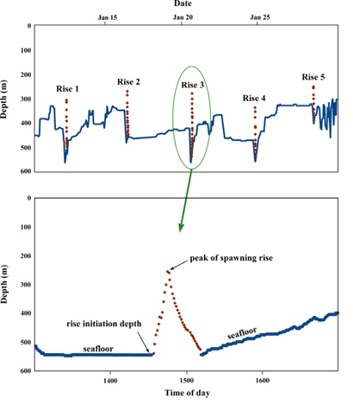
Figure 5. Upper panel: Depth profile of a Pacific halibut that conducted a series of five putative spawning rises during January, 2006. Based on the behavior of other flatfish, we hypothesize that reproductive products are released at the peak of each rise, with intervening periods composed of the hydration of subsequent batches of eggs, and potential courtship. Lower panel: The fish’s third spawning rise, enlarged. Data points are separated by one minute.
Taken together, the analysis of onshore-offshore migration and spawning rises can be used to characterize the spawning period for Pacific halibut, and study the degree to which spawning periods might vary among regions or change over time. The most comprehensive analysis was undertaken for fish that were tagged in the eastern Gulf of Alaska (Loher 2011). The data were compared with historical surveys that evaluated spawning condition of female Pacific halibut over the course of the winter (St. Pierre 1984) to produce an understanding of the average change in depth of likely females over the course of a year; the proportion of those individuals likely to have completed migrations to and from their spawning grounds at any given time; and the period over which the fish were actively spawning, defined as the period during which eggs are released into the water and fertilized. The general observations include: gradual movement of fish from the continental shelf to the continental slope that begins in late September and continues until January; return to the shelf that begins in January and continues through May; and an active spawning period that is concentrated from late December to early March. It is cautioned, however, that this analysis was based on a relatively small sample size (as few as 29 fish during the spring migration) and that the lengths of the fish that were tagged did not fully represent the exploitable stock. Furthermore, unpublished data suggest that migratory dynamics may depend to some extent upon the region in which the fish reside. In particular, data collected in the Bering Sea (Loher and Clark 2010) indicate regional patterns in both the timing and duration of migration to offshore waters.
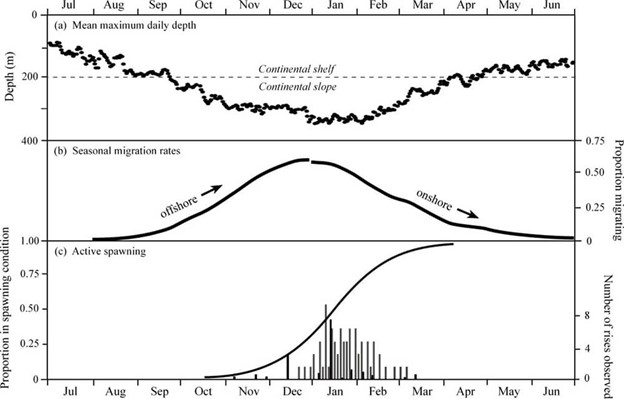
Figure 6. Composite depiction of seasonal movement and spawning in Pacific halibut in the eastern Gulf of Alaska: (a) Mean maximum depths occupied by Pacific halibut displaying seasonal onshore-offshore migration. (b) The proportion of the tagged population having initiated their fall offshore migration (left curve; n = 104) and completed their spring onshore migration (right curve; n = 29) over time. (c) Logistic fit of the proportion of reproductive fish sampled during historical surveys that were in spent condition (solid line), proportion of running fish in the same surveys (solid bars), and frequency of spawning rises observed in electronic tagging data (grey bars).